Endoplasmic Reticulum/Golgi Complex/Lysosomes
I. General Observations
Endomembrane
system (ES).
The recognized components of the ES (except for mitochondria, which are not
part of the ES) are shown in the following table:
| Intracellular Compartment | Percent of Total Cell Volume | Percent of Total Cell Membrane |
| Cytosol | 54 | 2 |
| Mitochondria | 22 | 7 (OM) + 32 (IM) |
| rough endoplasmic reticulum | 9 | 35 |
| smooth ER, plus Golgi Complex | 6 | 23 |
| nucleus | 6 | 0.2 |
| peroxisomes | 1 | 0.4 |
| lysosomes | 1 | 0.4 |
| endosomes | 1 | 0.4 |
The cytoplasmic face of ES membrane is directly opposed to the cytoplasm and
is similar
in composition to the inner face of the plasma membrane and the outer
faces of other components of the ES;
II. Structure of ER and Golgi Complex
A. ER
1. SER
2. RER
B. Golgi Complex (Dictyosomes for plants)
1. three membrane
components seen with the EM
2. Membrane differentiation in the Golgi Complex
The thickness of membranes increases progressively from the ER (or nuclear
envelope to the Golgi and plasma membrane:
| Membrane Type | Membrane Thickness (nm) |
| Nuclear envelope | 56 |
| ER | 53 |
| Golgi Complex cisternae 1 | 53 |
| 2 | 60 |
| 3 | 65 |
| 4&5 | 75 |
| Secretory Vesicles | 88 |
| Plasma membrane | 93 |
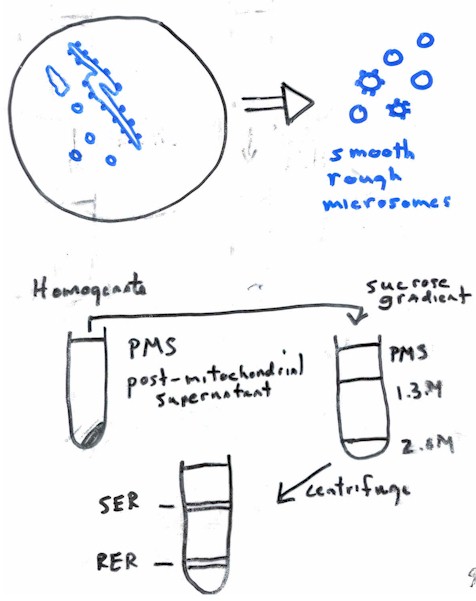
III. Isolation and Fractionation
A. General
comment: microsomes;
B. ER Isolation and fractionation
C. Isolation of Golgi Complex
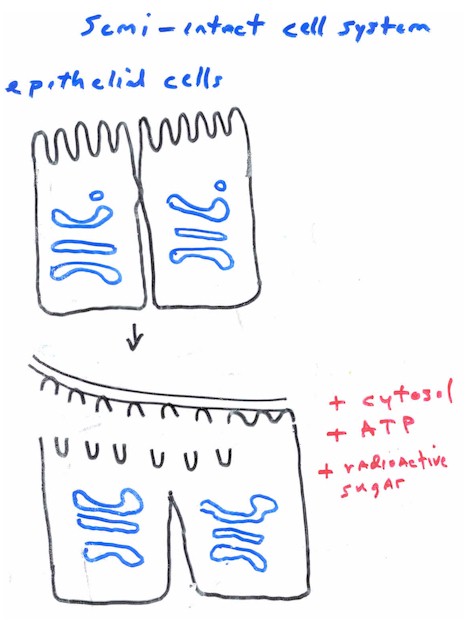
1. cell-free system
2. Semi-intact cell systems
IV. Enzyme Complement and Functions of the ER & Golgi Complex
A. ER
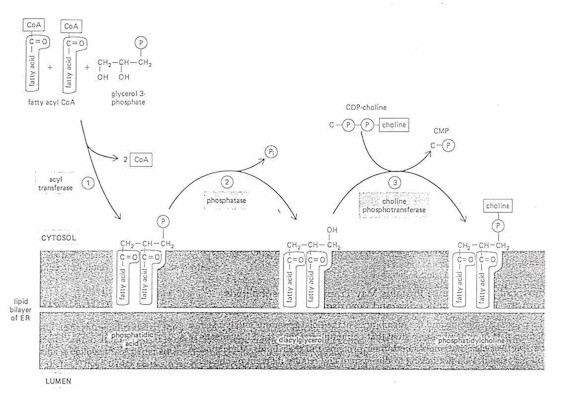
1. Membrane biogenesis
example: phosphatidyl choline (lecithin) - most common phospholipid (PL)
a. acyl transferase
glycerol 3 phosphate + fatty acid (in cytosol)---> phosphatidic acid
(fatty acids inserted on the cytosol side of the ER bilayer)
b. phosphatase
phosphatidic acid---> diacylglycerol + Pi
c. cholinephosphotransferase
diacylglycerol + CDP-choline---> phosphatidyl choline
Why does the membrane not become imbalanced with more phospholipid on the
cytosolic side?
2. Detoxification
a. Drug inactivation.
b. aryl hydroxylase
example reaction:
3-benzopyrene---> 5,6 epoxide
Where is 3-benzopyrene found?
3. Glycogenosis
in fasted lab animals, residual glycogen in the liver is often associated
with SER vesicles; the ER membrane contains the enzymes, glycogen phosphorylase which
breaks down glycogen into glucose-1-phosphate and glucose-6-phosphatase which breaks down
glucose-6-phosphate down into glucose. The conversion of glucose-1-phosphate to
glucose-6-phosphate is accomplished in the cytoplasm. The resulting glucose then can leave
the liver cell for export to the blood stream.
4. Calcium sequestration
5. ER and the Synthesis of Proteins
Questions:
Signal Hypothesis
This was proposed as a consequence of a study of secreted protein
synthesis using a cell-free (in vitro) system;
What is an example of a secreted protein?
What are "cell-free" systems?
It was found that if microsomes were omitted from the cell-free system,
proteins were larger (had more amino acids) than if the microsomes were made available to
the reaction;
What else was in the reaction vessel?
The increase in size of the protein was found to be because of the presence
of an amino terminal leader peptide; this leader peptide is synthesized first and directs
the translational complex to the ER; this leader peptide is then later cleaved off by an
enzyme in the ER membrane called signal peptidase.
More detail about the Signal Hypothesis
The process is somewhat more complicated in that the attachment of the signal
peptide to the ER membrane is guided by two other components:
SRP binds to the signal peptidase as soon as the protein emerges from the ribosome; this causes a pause in protein synthesis so that the ribosome can bind to the ER membrane
and not inappropriately release the protein into the cytoplasm;
translation resumes when the SRP binds to the docking protein which is found
on the cytosolic surface of the RER;
"co-translational translocation"
" post-translational translocation"
6. ER and Core Glycoslyation
The addition of sugars to proteins is one of the major functions of ER
(glycoprotein synthesis).
What is the overall function of a glycoprotein?
Most glycoproteins are eventually transported to the Golgi Complex, plasma
membrane and the lysosomes or peroxisomes.
The protein glycosylation process
This process involves the transfer of an oligosaccharide to a protein.
example: oligosaccharide with 14 sugar residues (n-acetyl glucosamine,
mannose, glucose);
oligosaccharide is always transferred to the NH2 group on side
chain of asparagine residue of the protein; thus, oligosaccharide is said to be "N"
linked;
dolichol is a long hydrophobic lipid with 22 five carbon units that span the
ER lipid bilayer three times;
The transfer of the oligosaccharide to asparagine is catalyzed by a membrane bound enzyme called glycosyl transferase;
the process
(image from http://www.colorado.edu/MCDB/MCDB1150/lectures/figure/dolichol.gif)
The diversity of glycoproteins comes from additional modification of N-linked
oligosaccharides and additional glycosyl transferases; these enzymes are found mostly in
the Golgi Complex.
B. Golgi Complex
Review
The Golgi Complex has four functionally distinct components: a cis or
entry face, medial cisterne, trans cisternae and the trans Golgi network (TGN)
How do materials move
through the Golgi Cisternae?
Cisternae of the Golgi are stable entities held together by a protein
scaffold. Material moves through Golgi cisternae by specific targeted vesicles that bud
from one compartment and fuse with the next. These vesicles are 'coated' with a
non-clathrin type protein to help form the vesicle.
How do proteins normally present in the ER avoid budding off and fusing
with the cis face of the Golgi Complex?
Resident proteins of the ER contain a sequence of amino acids
(lys-asp-glu-leu) which is lacking in Golgi proteins; a receptor for this sequence on the
internal surface of the ER keeps those proteins in place (some recycling also takes
place).
1. Glycoprotein processing
Remember that a single species of N-linked oligosaccharide is attached to
many different proteins in the ER; further processing occurs in the Golgi Complex;
stepwise additions of additional sugar residues to the core region are
catalyzed by other glycosyl transferases;
example: UDP-N-acetylglucosamine subunits are transferred into the Golgi by a
transmembrane carrier protein; the sugar is then attached by N-acetylglucosamine
transferase.
What is the purpose of N-linked glycosylation?
2. Proteolytic processing and cell secretion
Example: Insulin processing and secretion
Insulin is a hormone produced by the beta cells of the pancreatic islands; MW = 12,000; 2 polypeptide chains: A - 21 amino acids; B - 30 amino acids; A is linked to B with 2 disulfide bonds.
3. Vesicle targeting from the Golgi Complex to the cell surface
Vesicles produced in the ES must "know" which compartments to fuse
with; this recognition based upon the presence of a unique "molecular address
label" (at least ten are known) on the membrane surface that allows for the delivery
of its contents only to a specific membrane.
Two examples of vesicle targeting
a. Secretion
b. Mannose-6-phosphate receptor targeting to lysosomes
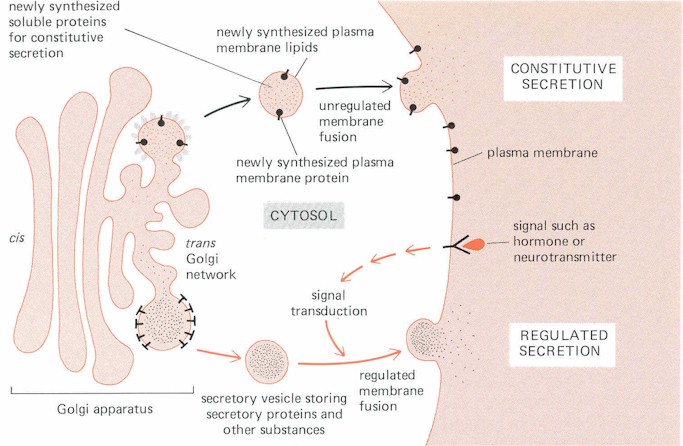
a. Secretion
Transport vesicles designed for immediate fusion with the plasma membrane
normally leave the Golgi Complex in a steady stream: membrane vesicle fuses with the PM
and then empties its contents into the extracellular fluid.
Two types of secretion
Constitutive pathway (vesicles immediately fuse with the PM);
Regulated pathway (vesicles are stored in the cytoplasm for later release in
response to some signal).
Note: secretory vesicles form by a budding process from the trans Golgi
network; the budding process is aided by the presence of clathrin
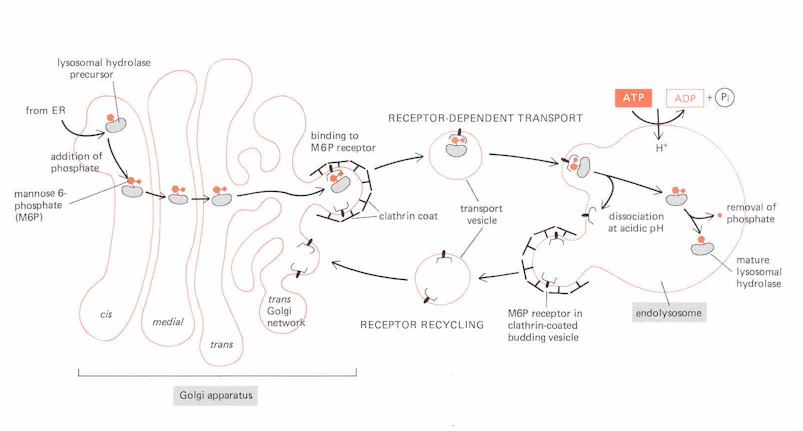
b. Mannose-6-phosphate receptor targeting to lysosomes
Review of the lysosome
Lysosomal hydrolases carry mannose-6-phosphate groups
attached to the N-linked oligosaccharide; complementary M6P receptors are found in the
membranes of clathrin coated
vesicles;
M6P receptors are transmembrane proteins which bind selectively to enzymes with M6P; this loads the Golgi vesicles prior to fusion with the endolysosome; the receptors are then recycled back to the trans-Golgi network by other transport vesicles.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}