| Adenylyl Cyclase and its role in Signal Transduction:
A Minireview
Sean Necessary, the University of Central Arkansas Adenylyl cyclase is a lyase class enzyme that
catalyzes the formation of cyclic adenosine monophosphate from adenosine
triphosphate. Adenylyl cyclase exists in three classes (I, II, and III)
and has nine isoforms in class III mammalian adenylyl cyclase. It exists
mainly in a membrane associated form but can also be found in a soluble
form depending on the organism or tissue. Adenylyl cyclase is a major player
in maintaining signal transduction pathways by converting numerous extracellular
hormonal signals into the form of a second messenger in cyclic adenosine
monophosphate. The active site, as well as some regulatory sites, is formed
as a result of dimerization of the enzymes two catalytic domains, which
is essential for catalysis. Regulation of adenylyl cyclase is performed
by activated G-proteins, calmodulin, P-site analog inhibitors, phosphorylation,
and forskolin, a hypotensive diterpene drug, at differing regulatory sites.
Two Mg2+ or Mn2+ ions are required to bind with adenosine triphosphate
in order for catalysis to occur. The production and regulation of cyclic
adenosine monophosphate is critically important in the activation of several
cellular metabolic processes, including enzyme activity, gene-expression
patterns, and membrane excitability, making correct adenylyl cyclase function
essential in the maintenance of normal physiology.
Introduction:
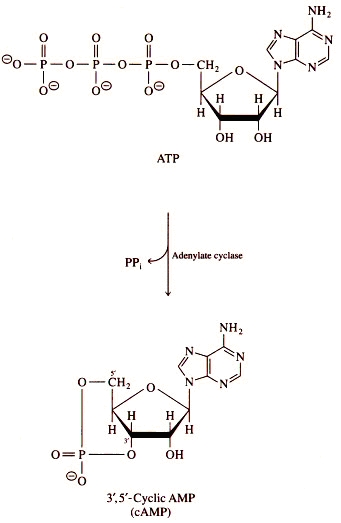
Figure 1. Overall adenylyl cyclase catalyzed reaction of cAMP and pyrophosphate (PPi) from ATP (3). also known by other common names such as adenylate
cyclase and 3',5'-cAMP synthetase (5).
AC exists in most cells as a transmembrane protein in small amounts less
than 10 pmols/mg of membrane, but is also found in soluble form in the
cytoplasm of some cell types and unicellular organisms (2,6,7).
However, mammalian transmembrane AC and soluble AC are suspected to have
evolved separately based upon the greater similarities of portions of the
catalytic domains of sAC with those of bacterial AC than with transmembrane
AC (8).
AC acts as a signal converter as it receives signals from hormones and
other molecules that ultimately activate it to catalyze the formation of
3',5'-cAMP, an important second messenger in signal transduction pathways
(9). The signal that AC helps to transduce comes from various external
hormonal stimuli that bind to and activate membrane associated G-proteins.
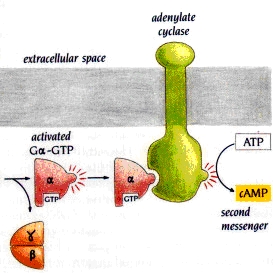
Figure 2. Gsa activation of adenylyl cyclase, increasing the intracellular concentration of cAMP (3). Structure:
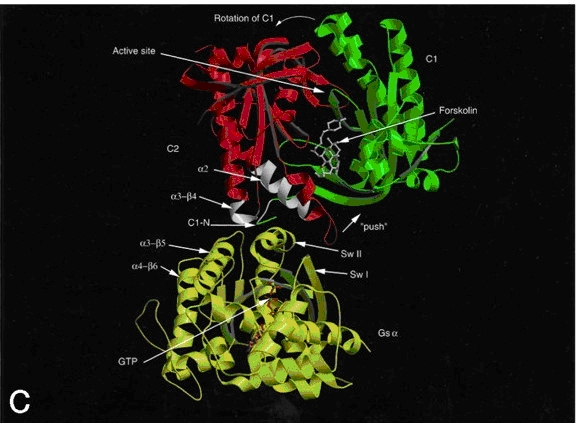
Figure 3. Ribbon diagram of AC with forskolin bound to its allosteric site and the Gsa allosteric site colored white. Gsa is activated with GTP bound. The green and red domains are C1 and C2, respectively. The ATP binding site is just above the forskolin binding site. Not shown are the two transmembrane domains (10). Kinetics and Catalytic Mechanism:
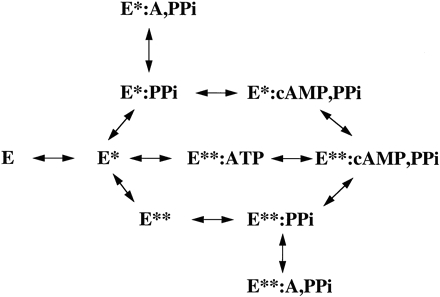
Figure 4. Schematic of enzyme cycle of AC, including enzyme conformation and substrate/products bound (10). Within the active site, residues Asp354,
Arg398, Lys938, Arg1011, Asp1018, Asn1025, Arg1029, and Lys1067 are
involved in ATP-Mg2+/Mn2+ binding and catalysis (3,10).
These residues are brought into correct position by a 7° rotation of
the C1/C2 domains with respect to each other during activator induced dimerization.
The adenine ring of ATP binds to a hydrophobic pocket within the active
site and is also stabilized by water mediated interaction between the polar
amine group and Asn1025. However, the greatest specificity for adenine
is determined by hydrogen bonding between the Asp1018 and the adenine N6,
as well as hydrogen bonding of Lys938 and the adenine N1. These two residues,
Asp1018 and Lys938, differentiate AC from being a GTP binding guanyl cyclase
(GC) (10,16).
Diseases:
Figure 5. Diagram of the hormonal stimulation of different G-protein receptors and the up or downregulation of cAMP production. cAMP is shown activating protein kinase A (PKA), which regulates multiple cellular metabolic functions (7). The functions affected depend upon the hormonal
stimulus and the hormones target tissue. For example, when the hormone
adrenaline binds to G-protein associated receptors in cells of the heart,
cAMP is upregulated by the activation of AC. cAMP then stimulates an increase
in heart rate and contraction strength. The same hormonal stimulus targets
muscle and fat cells, but causes different responses. In muscle, cAMP induces
glycogen breakdown, and in fat cells, cAMP induces lypolysis. Thus, the
abnormal production or regulation of cAMP could have major detrimental
effects on these metabolic processes. G-proteins are regulated by numerous
other hormones including glucagon, secretin, and parathyroid, among others,
which all confer induction of separate signal transduction cascades. However,
all theses cascades are sustained by only a few forms of AC within an organism,
making ACs function vitally important in cellular metabolic function (7,9,10,17).
This makes implications of disease and metabolic abnormalities associated
with AC dysfunction due to mutations or disease almost endless. Examples
of disease caused by AC dysfunction or dysfunction in AC regulation include
whooping cough and malignant hyperthermia. Whooping cough is caused by
the pertussis toxin produced by the bacterium Bordetella pertussis. Ultimately
pertussis toxin blocks inhibition of AC by Gia leading to an overproduction
of cAMP in multiple tissues. This overproduction disrupts normal functions
regulated by cAMP, conferring the diseased state, in this case, whooping
cough (17).
Malignant hyperthermia (MH) is an example of the effects of high AC activity.
MH results from a high cAMP concentration in skeletal muscle that accumulates
as a result of abnormally elevated AC activity (18).
References:
2. Barman, T. E. (1974) Enzyme Handbook: Supplement I, p. 457, Springer-Verlag New York Heidelberg Berlin 3. Robbins, M. (2000), Tutorial: Adenylyl Cyclase, BIMCORE <http://www.bimcore. emory.edu/home/Kins/bimcoretutorials/Mrobbin/bioweb/adenylyl/text.html> 4. Adenylate Cyclase (2005) Wikipedia: The Free Encyclopedia, Wikimedia Foundation, Inc. <http://en.wikipedia.org/wiki/Adenylate _cyclase> 5. Reshma (2005), Human Protein Reference Database- Adenylyl Cyclase Soluble, Institute of Bioinformatics, Bangalore, India <http://www.hprd.org/protein/05549?selectedtab=ALTERNATE+NAMES> 6. Hammes, G. (1982) Enzyme Catalysis and Regulation, Academic Press Inc. 7. Other Inhibitors: Adenylate Cyclase Inhibitors, EMD Biosciences (2006) <http://www.emdbiosciences.com/html/CBC/other_inhibitors_adenylate_cyclase.htm> 8. Buck, J., Cann, M. J., Levin, L. R., Schapal, L., Sinclair, M. L. (1999), Cytosolic adenylyl cyclase defines a unique signaling molecule in mammals, Proc Natl Acad Sci U S A., 96(1): 7984. <http://www.pubmedcentral.gov/articlerender.fcgi?tool=pubmed&pubmedid=9874775> 9. Berg, J. M., Tymoczko, J. L., Stryer, L. (2002) Biochemistry, 5th Ed., Ch. 15, W. H. Freeman and Company 10. Hurley, J. H., Tang, Wei-Jen (1998), Catalytic Mechanism and Regulation of Mammalian Adenylyl Cyclases, Molecular Pharmacology, 54, 231-240 <http://molpharm.aspetjournals.org/cgi/content/full/54/2/231> 11. Milligan, G., Wakelam, M. (1992) G-Proteins: Signal Transduction & Disease, Ch. 1 & 2, Academic Press Limited 12. Alberts, B., Bray, D., Hopkin, K., Johnson, A., Lewis, J., Raff, M., Roberts, K., Walter, P. (2004) Essential Cell Biology, 2nd Ed., Garland Science 13. IPR000274 Adenylate cyclase, class-I (2005-06), European Molecular Biology Laboratory- European Bioinformatics Institute (EMBL- EBI) <http://www. ebi.ac.uk/interpro/IEntry?ac=IPR000274> 14. Bowen, R. (2000) Adenylyl Cyclase,<http://arbl.cvmbs.colostate.edu/hbooks/molecules/cyclase.html> 15. Forskolin (2000), WholeHealthMD.com, LLC. <http://www.wholehealthmd.com/ refshelf/substances_view/0,1525,10025,00. html> 16. Sunahara Lab: Adenylyl Cyclase (2006), University of Michigan. <http://calliope. med.umich.edu/Alternative%20Website/Sunahara-home1.html> 17. Todar, K. (2004), Bordetella pertussis and Whooping Cough, Univ. of Wisconsin-Madison Dept. of Bact. <http:// textbookofbacteriology.net/pertussis.html> 18. Cerri, C. G., Willner, J. H., and Wood, D.
S. (1981), High skeletal muscle adenylate cyclase in malignant hyperthermia,
J Clin Invest. 68(5): 11191124. <http://www.pubmedcentral.com/articlerender.fcgi?artid=370904>
|