 |
 |
 |
|
|
|
|
 |
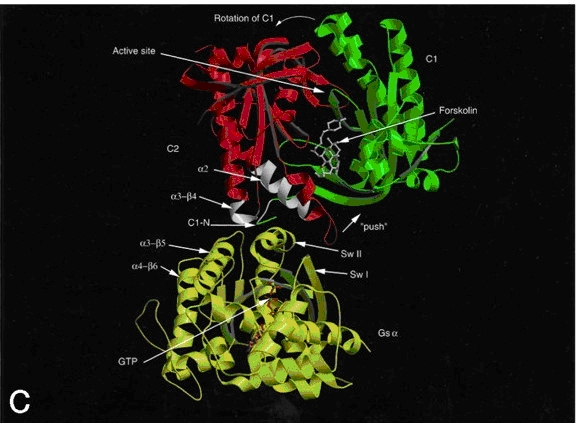 Figure 3. Ribbon diagram of AC with forskolin bound to its allosteric site and the Gsa allosteric site colored white. Gsa is activated with GTP bound. The green and red domains are C1 and C2, respectively. The ATP binding site is just above the forskolin binding site. Not shown are the two transmembrane domains. |
There are three classes of AC,
however, the structure of the active site is highly conserved across all
classes. Class I-ACs are found in Gram negative bacteria such as E coli.
Class II-ACs, or toxic AC, are found in pathogenic microbes such as Bordetella
pertussis and Bacillus anthracis. Finally, class III-ACs are found in a
wide range of organisms, including nine isoforms, AC-1 through AC-9, found
in mammals . AC has cytoplasmic soluble (sAC) forms and membrane associated
forms. The membrane associated forms consist of two transmembrane domains
of 6 a-helices each. Both the soluble and membrane associated forms have
two highly conserved cytoplasm associated catalytic domains, C1 and C2.
In the membrane associated forms, C1 bridges the two transmembrane domains
and C2 makes up the C-terminal end of the protein. C1 and C2 come together
to form separate active and regulatory sites at their interface.
Regions of C1 and C2, namely C1a and C2a, have been shown through mutational analysis to contribute the amino acids that make up the ATP binding and active site. The regulatory site at the interface of the joined catalytic domains is a potential binding site for ATP, but, in most mammalian isoforms, has a greater affinity for forskolin, a hypotensive diterpene usually administered to patients with hypothyroidism. Forskolin binds at a large hydrophobic pocket that lies proximal to the active site, thus, stabilizing the C1/ C2 dimer. Another, more significant, allosteric site lies approximately 30 Å from the catalytic site. This site also forms as a result of the C1/ C2 dimerization and is the site of Gsa binding. Thus, Gsa, along with forskolin, stabilize the C1/ C2 dimer, activating AC. Gsa is the GTP activated stimulatory alpha subunit of a heterotrimeric G-protein after it receives appropriate hormonal stimulus. The Gsa site is 1800 Å2 and is important in inducing an activating conformational change of AC (Fig. 3). Noncompetive inhibition of AC is accomplished by binding of Gia inhibitory subunit of Gi family of G-proteins to a currently unidentified regulatory site. Ca2+ and calmodulin have been shown to regulate adenylyl cyclase at regulatory site other than that of the G-protein and forskolin sites. Also phosphorylation, by protein kinase A (PKA) or PKC, of Ser674 in the C1b region reduces AC-VI activity by 50%. An analog of ATP, P-site inhibitor, shows a higher affinity within the active site of AC and has been used extensively to determine the kinetics and mechanism of AC. (see My Adenylyl Cyclase Minireview for more detail and referencing) |